Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
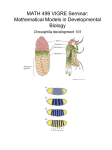
Multicellular development and morphogens Development is the successive process of systematic gene-directed changes throughout an organism’s life cycle -Can be divided into four sub-processes: -Growth (cell division) -Differentiation -Pattern formation -Morphogenesis Pattern formation: • the generation of complex organizations of cell fates (programming of a cell to follow a specified path of cell differentiation) in space and time. • Pattern formation is controlled by genes . Morphogen • Is a substance governing the pattern of tissue development and, in particular, the positions of the various specialized cell types within a tissue. • It spreads from a localized source and forms a concentration gradient across a developing tissue. Morphogenesis: • Is developmental process by which anatomical structures or cells shape and size are generated and organized. • It achieves through changes in: -Cell division -Cell shape and size -Cell death -Cell migration( animal) • Morphogenesis is controlled by certain types of molecules: 1) morphogens. 2) transcription factors. 3) cell adhesion molecules. • The morphogen idea has a long history in developmental biology, dating back to the work of the pioneering Drosophila geneticist, Thomas Hunt Morgan, in the early 20th century. • However, it was Lewis Wolpert who refined the morphogen concept in the 1960s with his famous French flag model which described how morphogen could subdivide a tissue into domains of different target gene expression (corresponding to the colours of the French flag). • In developmental biology a morphogen is rigorously used to mean a signaling molecule that acts directly on cells (not through serial induction) to produce specific cellular responses dependent on morphogen concentration. • During early development, morphogen gradients generate different cell types in distinct spatial order. • The morphogen provides spatial information by forming a concentration gradient that subdivides a field of cells by inducing or maintaining the expression of different target genes at distinct concentration thresholds. • Distinct cell types emerge as a consequence of the different combinations of target gene expression. • In this way, the field of cells is subdivided into different types according to their position relative to the source of the morphogen. • Cells far from the source of the morphogen will receive low levels of morphogen and express only lowthreshold target genes. • In contrast, cells close to the source of morphogen will receive high levels of morphogen and will express both low- and high-threshold target genes. Mutations of HNF-1 inhibit epithelial morphogenesis through dysregulation of SOCS-3 • Hepatocyte nuclear factor-1 (HNF-1) is a Pit-1, Oct-1/2, Unc-86 (POU) homeodomain-containing transcription factor expressed in the kidney, liver, pancreas, and other epithelial organs. • Mutations of HNF-1 cause maturity-onset diabetes of the young, type 5 (MODY5), which is characterized by early-onset diabetes mellitus and congenital malformations of the kidney, pancreas, and genital tract. • Knockout of HNF-1 in the mouse kidney results in cyst formation. Zhendong Ma*, Yimei Gong*, Vishal Patel*, Courtney M. Karner*†, Evelyne Fischer‡, Thomas Hiesberger*,Thomas J. Carroll*†, Marco Pontoglio‡, and Peter Igarashi*§¶Edited by Maurice B. Burg, National Institutes of Health, Bethesda, MD, and approved October 25, 2007 (received for review June 25, 2007) Role of CDMP-1 in Skeletal Morphogenesis: Promotion of Mesenchymal Cell Recruitment and Chondrocyte Differentiation Noriyuki Tsumaki,* Kazuhiro Tanaka,* Eri Arikawa-Hirasawa,* Takanobu Nakase, Tomoatsu Kimura,§ J. Terrig Thomas, Takahiro Ochi, Frank P. Luyten, and Yoshihiko Yamada* Cartilage-derived morphogenetic protein-1 (CDMP-1), a member of the bone morphogenetic protein family, it has role of in skeletal formation. Characterization of bone morphogenetic protein family members as neurotrophic factors for cultured sensory neurons L. M. Farkas, J. Jászai, K. Unsicker and K. Krieglstein Neuroanatomy, University of Heidelberg, Im Neuenheimer Feld 307, D-69120 Heidelberg, Germany • The neurotrophic factors may act synergistically in ensuring neuronal survival. • The bone morphogenetic proteins have been implicated in several inductive processes throughout vertebrate development including nervous system patterning. • Several bone morphogenetic proteins can be detected in developing embryonic day 14 rat dorsal root ganglia . • Growth/differentiation factor-5, bone morphogenetic protein-2, -4, -7 and -12 significantly increased the survival promoting effects of neurotrophin-3 and nerve growth factor on cultured dorsal root ganglion neurons. Failure of bone morphogenetic protein receptor trafficking in pulmonary arterial hypertension Anastasia Sobolewski1, Nung Rudarakanchana1, Paul D. Upton1, Jun Yang1, Trina K. Crilley1, Richard C. Trembath2 and Nicholas W. Morrell1,* • Heterozygous germline mutations in the gene encoding the bone morphogenetic protein type II receptor cause familial pulmonary arterial hypertension (PAH). • substitution of cysteine residues in the ligand-binding domain of this receptor prevents receptor trafficking to the cell membrane. • HeLa cells were transiently transfected with BMPR-II wild type or mutant (C118W) receptor constructs. Bone Morphogenetic Protein Receptor2 Mutations in Pulmonary Hypertension Jane H. Morse Chest 2002;121;50S-53S Heterozygous mutations in bone morphogenetic protein receptor (BMPR) 2 cause PPH Mutations in the Gene Encoding Capillary Morphogenesis Protein 2 Cause Juvenile Hyaline Fibromatosis and Infantile Systemic Hyalinosis Sandra Hanks1,Sarah Adams1,Jenny Douglas1,Laura Arbour2,David J. Atherton3,Sevim Balci7,Harald Bode8,Mary E. Campbell4,Murray Feingold9,Gökhan Keser10,Wim Kleijer11,Grazia Mancini11,John A. McGrath5,Francesco Muntoni6,Arti Nanda12,M. Dawn Teare13,Matthew Warman14,F. Michael Pope4,Andrea Superti-Furga15,P. Andrew Futreal16andNazneen Rahman1,, • Juvenile hyaline fibromatosis (JHF) and infantile systemic hyalinosis (ISH) are autosomal recessive conditions characterized by multiple subcutaneous skin nodules, gingival hypertrophy, joint contractures, and hyaline deposition. Developmental genes: Two types: 1.maternal effect genes: affects early development through contributions of gene products from the ovary of the mother to the developing oocyte. a) genes regulate anterior-posterior axis pattering: bicoid –hunchback-caudal-nanos gene. b) genes regulate dorsal-ventral axis pattering. dorsal gene (dl). • Zygotically acting genes: are those in which gene products contributing to early development are expressed exclusively in the zygote. • For example: a) segmentation genes b) homeotic genes. Maternal effect gene : • Maternal effects often occur because the mother supplies particular mRNA to the oocyte. • The phenotype is thus determined by the mother's, rather than the egg's, genotype. • The oocyte is formed, along with fifteen nurse cells, from a stem cell. • The nurse cells secrete maternal mRNAs into the developing oocyte through a structure known as ring canal. OOGENESIS IN DROSOPHILA Germarium Germline Cyst Formation Cystoblast germline stem cells follicle stem cells ring canal Pro-Oocyte (undergoes meiosis) germline: stem cell > cystoblast > 1 oocyte + 15 nurse cells Class of maternal effect genes: • Bicoid gene: key maternal effect gene involved in the formation of the anterior structure of the egg. • Bicoid protein diffuse to form a gradient with it’s highest concentration at the anterior end of the egg it’s direct the formation of head and thorax. • It act as transcription activator and translation repressor. Nanos genes: • Key maternal effect gene involved in the formation of the posterior structures of the embryo. • Nanos or (nos) protein forms a posterior-anterior gradient. • Directs the formation of the abdomen. • Nanos protein is a translational repressor (direct the concentration of hunchback mRNAs). • There are specific microtubule- association sequences located within the 3’ UTRs of the mRNA. • Those sequences are bound by a protein that can also bind the – ends of the microtubule in case of bcd mRNA and + end in case of the nanos mRNA. Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Maturation Of Drosophila Oocyte Movement of bicoid mRNA moves maternal mRNA toward anterior end Follicle cells Nurse cells Anterior Posterior Microtubules nanos mRNA moves toward posterior end a. Nucleus Anterior Posterior bicoid mRNA b. nanos mRNA Caudal genes & maternal hunchback genes: • Cad mRNAs and hunchback mRNAs are evenly distributed in the egg before fertilization because they lack special location control system. • CAUDAL protein has a lowest concentration at the anterior end and highest concentration at the posterior end. • It activate genes needed for the formation of the posterior end. • HUNCHBACK protein has a highest concenyration at the anterior end and it’s responsible for the formation of the anterior end. • Bicoid protein gradient forms at the anterior end of the egg. • Nanos protein forms a gradient at the posterior end. • The Bicoid protein blocks translation of caudal mRNA so Caudal protein is made only in the posterior part of the cell. • Nanos protein binds to the hunchback mRNA and blocks its translation in the posterior end. Control of hunchback mRNA Translation by Nanos Protein: Three independent Genetic Pathways Interact to Form th Anterior-Posterior Axis of the Drosophila Embryo Experiments Demonstrating That the Bicoid Gene Encodes the Morphogen Responsible for Head Structures in Drosophila Experiments Demonstrating that the Bicoid Gene Encodes the Morphogen Responsible for Head Structures in Drosophila Segmentation genes • The segmentation genes give 14 parasegments that are closely related to the final anatomical segments. • Mutations in segmentation genes alter the number of segments or the internal organization. • Gap Genes • Pair-rule Genes • Segment Polarity Genes Gap genes • The gap genes are the first of a cascade of the segmentation control genes. • These genes map out the basic subdivisions along the anteriorposterior axis. establish broad regional domains of body plan of the embryo each of which covers areas that will later develop into several distinct segments. • All encode transcriptional factors. • Mutation in gab genes results in the deletion of regions consisting of several adjusting segments. • The gradients of Maternal effect (Bicoid, and Caudal) work as transcriptional regulation (activators/ repressor) for Gap gene expression. • Bicoid activates Kruppel and hunchback and others; their proteins spread and activate pair-rule genes, etc. Sequential subdivision strategy. • The resultant proteins from these genes become stabilized and maintained by interactions between the different gap genes themselves. • activity is characterized by low levels of transcriptional activity across the entire embryo, with discreet areas of high activity forming as cleavage continues. • Transcription patterns of anterior gap genes are inititated by the different concentrations of Hunchback and Bicoid. • in the anterior of the embryo high Hunchback leads to the transcription of Giant. • lower Hunchback leads to the transcription of Krüppel represses posterior expression Giant and Knirps . • knirps inhibits hunchback transcription, thereby setting the posterior boundary of Hunchback. • Caudal protein which is highest gradient in the posterior portion of the embryo activate knirps and giant. Gt: gaint Hb: hunchback Kr: kruppl Kni: knirp pair-rule genes • pair-rule genes expression leads to division of the embryo into number of regions, each of which contains a pair of parasegments. cover up a pattern of 7 repeats vertical to the anterior-posterior axis, then subdivide to 14 repeats. • All encode transcriptional factors. They are controlled directly by the gap gene proteins, which activate the transcription of some pair-rule genes while repressing the transcription of others. • Mutations result in embryo with half the normal segment number either odd- or evennumbered stripes. • Mutant at fushi-tarazu gene • gap genes initiate the expression of members of the pair-rule class )even-skipped, hairy ,and runt( primary" pair-rule genes. • Interactions between primary pair-rule genes become self-stabilized once inititated, and they are responsible for activation or inhibiting the secondary pair-rule genes. • Pair-rule and gap genes also interact to regulate the homeotic genes that determine the identity of each segment. • Interestingly, although the pair-rule genes were identified genetically by their pattern defects in alternate segments, this class of segmentation genes is expressed in a wide variety of tissues during embryogenesis. • pair-rule genes are expressed in the mesoderm, gut and most notably the central nervous system . polarity genes • set the anterior-posterior axis of each segment that will become the segments seen in the larva and adults and responsible for maintaining certain repeating structures within the segments. • Mutations produce embryos with the normal segment number, but with part of each segment replaced by a mirrorimage. • The majority are transcriptional factor but are also signaling protein as hedgehog. • This expression pattern is initiated by the pair-rule genes (like even-skipped) that code for transcription factors that regulate the other genes. • Mutations results in segments replaced by mirror images . • Many segment polarity genes encode proteins that are part of the Hedgehog and Wnt cellsignalling pathways (ligands, transmembrane proteins, receptors, signal transducers, transcription factors). Homeotic genes The final stage of specification of segment body plan is established by activation of genes called homeotic selector genes These genes are responsible for controlling the developmental program that that determine the final cell fat decision of each segments The importance of their function is obviously clear in their mutant phenotype Homeotic genes Homeo refer to transformation of one structure or segment to another. There are 8 genes identified as homeotic selector gene Each one code for transcription factor that contain DNA binding motif (hoeodomain) The 8 genes are collectively organized in a complex known as Hom-c Homeotic genes Where they are divided into 2 groups: ANTENNEPEDIA complex which include : Antennapedia (Antp) Labial (lab) Proboscidea (pb) Deformed (dfd) Sex combs (scr) The genes of this cmplex involved in the specification of head and thoracic segment. Homeotic genes The bithorax complex which involve: abdominal-B ultrabithorax abdominal-A abdominal-B Which are involved in the specification of abdominal segment identities. There is a striking correlation between the organization of the homeotic selector genes within the fly genome and their transcription expression domains along A-P axis of the embryo and the adult fly. The linear arrangement of the genes in the HOM-C complex correlate precisely with their expression along the A-P axis. Homeotic genes The expression of the homeotic genes require in put from gab genes and pair-rule genes that control their expression within parasegmental domain. Gap and pair-rule protein act through cis- acting regulatory region (initiator enhancer elements to their activator or repress homeotic gene expression. For example the graidient of Hunchback and Krupple negatively regulate the expression of abdA, abd B in the head and thorcic segment. Another control mechanism : as transcription factor the protein the protein coded by the homeotic selector gene act to regulate the expression of each other. Homeotic genes For example the expression of Antennapidia complex is repressed by the combined activity of the bithorx complex protein. The absence of Ubx cause the posterior part of the Antennapidia to expand into a region normally express ubx. The homeotic selector genes are required for segment identity during larval and adult stage , their for another mechanism are needed because their regulation (gab,pair-rule) during embryo is transient How do the drosophilla maintain their homeotic genea expression during late stage? Homeotic genes The modification of chromatin structre play a major role in their controlling mechanism in adult which is carried out by two classes of genes: Polycomb group: This group inactivate the cis-regulatory region that control homeotic gene expression ,by remodeling chromatin structure into more compact states threr by inhibit their transcription during subsequent stages. Trithorax group: There are protein appear to act collectively to keep chromatin in a state that favors transcriptional activation. Homeotic genes Homeotic genes are responsible for regulating the specific genetic program that direct cells within each segments toward their final cell fats? Their expression is not restricted to an individual segment , most are expressed in broader pattern along the A-P axis All homeotic selector genes share highly conserved DNA-binding domain. ? This contribute to weak binding specificity for an individual homeotic genes to a particular region than other.? They depend on cofasctor proteins modulsate their activity in a context- dependent manner