Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
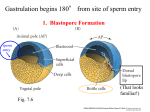
LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 47 Animal Development Lectures by Erin Barley Kathleen Fitzpatrick © 2011 Pearson Education, Inc. Overview: A Body-Building Plan • A human embryo at about 7 weeks after conception shows development of distinctive features 1 mm © 2011 Pearson Education, Inc. Concept 47.1: Fertilization and cleavage initiate embryonic development • Fertilization is the formation of a diploid zygote from a haploid egg and sperm © 2011 Pearson Education, Inc. The Acrosomal Reaction • The acrosomal reaction is triggered when the sperm meets the egg • The acrosome at the tip of the sperm releases hydrolytic enzymes that digest material surrounding the egg © 2011 Pearson Education, Inc. Figure 47.3-1 Basal body (centriole) Sperm head Acrosome Jelly coat Sperm-binding receptors Vitelline layer Egg plasma membrane Figure 47.3-2 Basal body (centriole) Sperm head Acrosome Jelly coat Sperm-binding receptors Hydrolytic enzymes Vitelline layer Egg plasma membrane Figure 47.3-3 Sperm nucleus Basal body (centriole) Sperm head Acrosome Jelly coat Sperm-binding receptors Acrosomal process Actin filament Hydrolytic enzymes Vitelline layer Egg plasma membrane Figure 47.3-4 Sperm plasma membrane Sperm nucleus Basal body (centriole) Sperm head Acrosome Jelly coat Sperm-binding receptors Acrosomal process Actin filament Fused plasma membranes Hydrolytic enzymes Vitelline layer Egg plasma membrane Figure 47.3-5 Sperm plasma membrane Sperm nucleus Basal body (centriole) Sperm head Acrosome Jelly coat Sperm-binding receptors Fertilization envelope Acrosomal process Actin filament Cortical Fused granule plasma membranes Hydrolytic enzymes Perivitelline space Vitelline layer Egg plasma membrane EGG CYTOPLASM • Gamete contact and/or fusion depolarizes the egg cell membrane and sets up a fast block to polyspermy © 2011 Pearson Education, Inc. The Cortical Reaction • Fusion of egg and sperm also initiates the cortical reaction • Seconds after the sperm binds to the egg, vesicles just beneath the egg plasma membrane release their contents and form a fertilization envelope • The fertilization envelope acts as the slow block to polyspermy © 2011 Pearson Education, Inc. • The cortical reaction requires a high concentration of Ca2 ions in the egg • The reaction is triggered by a change in Ca2 concentration • Ca2 spread across the egg correlates with the appearance of the fertilization envelope © 2011 Pearson Education, Inc. Fertilization in Mammals • Fertilization in mammals and other terrestrial animals is internal • Secretions in the mammalian female reproductive tract alter sperm motility and structure • This is called capacitation and must occur before sperm are able to fertilize an egg © 2011 Pearson Education, Inc. • Sperm travel through an outer layer of cells to reach the zona pellucida, the extracellular matrix of the egg • When the sperm binds a receptor in the zona pellucida, it triggers a slow block to polyspermy • No fast block to polyspermy has been identified in mammals © 2011 Pearson Education, Inc. Figure 47.5 Zona pellucida Follicle cell Sperm basal body Sperm nucleus Cortical granules Cleavage • Fertilization is followed by cleavage, a period of rapid cell division without growth • Cleavage partitions the cytoplasm of one large cell into many smaller cells called blastomeres • The blastula is a ball of cells with a fluid-filled cavity called a blastocoel © 2011 Pearson Education, Inc. Figure 47.6 50 m (a) Fertilized egg (b) Four-cell stage (c) Early blastula (d) Later blastula Cleavage Patterns • In frogs and many other animals, the distribution of yolk (stored nutrients) is a key factor influencing the pattern of cleavage • The vegetal pole has more yolk; the animal pole has less yolk • The difference in yolk distribution results in animal and vegetal hemispheres that differ in appearance © 2011 Pearson Education, Inc. Figure 47.7 Zygote 2-cell stage forming Gray crescent 0.25 mm 8-cell stage (viewed from the animal pole) 4-cell stage forming 8-cell stage Animal pole 0.25 mm Blastula (at least 128 cells) Vegetal pole Blastula (cross section) Blastocoel Figure 47.7a-5 Animal pole Gray crescent Zygote 2-cell stage forming Blastocoel Vegetal pole 4-cell stage forming 8-cell stage Blastula (cross section) Concept 47.2: Morphogenesis in animals involves specific changes in cell shape, position, and survival • After cleavage, the rate of cell division slows and the normal cell cycle is restored • Morphogenesis, the process by which cells occupy their appropriate locations, involves – Gastrulation, the movement of cells from the blastula surface to the interior of the embryo – Organogenesis, the formation of organs © 2011 Pearson Education, Inc. • The three layers produced by gastrulation are called embryonic germ layers – The ectoderm forms the outer layer – The endoderm lines the digestive tract – The mesoderm partly fills the space between the endoderm and ectoderm • Each germ layer contributes to specific structures in the adult animal Video: Sea Urchin Embryonic Development © 2011 Pearson Education, Inc. Figure 47.8 ECTODERM (outer layer of embryo) • Epidermis of skin and its derivatives (including sweat glands, hair follicles) • Nervous and sensory systems • Pituitary gland, adrenal medulla • Jaws and teeth • Germ cells MESODERM (middle layer of embryo) • Skeletal and muscular systems • Circulatory and lymphatic systems • Excretory and reproductive systems (except germ cells) • Dermis of skin • Adrenal cortex ENDODERM (inner layer of embryo) • Epithelial lining of digestive tract and associated organs (liver, pancreas) • Epithelial lining of respiratory, excretory, and reproductive tracts and ducts • Thymus, thyroid, and parathyroid glands Gastrulation in Sea Urchins • Gastrulation begins at the vegetal pole of the blastula • Mesenchyme cells migrate into the blastocoel • The vegetal plate forms from the remaining cells of the vegetal pole and buckles inward through invagination © 2011 Pearson Education, Inc. • The newly formed cavity is called the archenteron • This opens through the blastopore, which will become the anus © 2011 Pearson Education, Inc. Figure 47.9 Animal pole Blastocoel Mesenchyme cells Vegetal plate Vegetal pole Blastocoel Filopodia Mesenchyme cells Blastopore Archenteron 50 m Blastocoel Ectoderm Key Future ectoderm Future mesoderm Future endoderm Mouth Mesenchyme (mesoderm forms future skeleton) Archenteron Blastopore Digestive tube (endoderm) Anus (from blastopore) Gastrulation in Frogs • Frog gastrulation begins when a group of cells on the dorsal side of the blastula begins to invaginate • This forms a crease along the region where the gray crescent formed • The part above the crease is called the dorsal lip of the blastopore © 2011 Pearson Education, Inc. Figure 47.10 1 CROSS SECTION SURFACE VIEW Animal pole Blastocoel Dorsal lip of blastopore Early Vegetal pole gastrula Blastopore Blastocoel shrinking 2 3 Blastocoel remnant Dorsal lip of blastopore Archenteron Ectoderm Mesoderm Endoderm Key Future ectoderm Future mesoderm Future endoderm Late gastrula Blastopore Blastopore Yolk plug Archenteron Figure 47.11 Fertilized egg Primitive streak Embryo Yolk Primitive streak Epiblast Future ectoderm Blastocoel Migrating cells (mesoderm) Endoderm Hypoblast YOLK Gastrulation in Humans • Human eggs have very little yolk • A blastocyst is the human equivalent of the blastula • The inner cell mass is a cluster of cells at one end of the blastocyst • The trophoblast is the outer epithelial layer of the blastocyst and does not contribute to the embryo, but instead initiates implantation © 2011 Pearson Education, Inc. Figure 47.12 1 Blastocyst reaches uterus. Uterus Endometrial epithelium (uterine lining) Inner cell mass Trophoblast Blastocoel 2 Blastocyst implants (7 days after fertilization). Expanding region of trophoblast Maternal blood vessel Epiblast Hypoblast Trophoblast 3 Extraembryonic membranes start to form (10–11 days), and gastrulation begins (13 days). Expanding region of trophoblast Amniotic cavity Epiblast Hypoblast Yolk sac (from hypoblast) Extraembryonic mesoderm cells (from epiblast) Chorion (from trophoblast) 4 Gastrulation has produced a three-layered embryo with four extraembryonic membranes. Amnion Chorion Ectoderm Mesoderm Endoderm Yolk sac Extraembryonic mesoderm Allantois Figure 47.12a Endometrial epithelium (uterine lining) Uterus Inner cell mass Trophoblast Blastocoel 1 Blastocyst reaches uterus. Figure 47.12b Expanding region of trophoblast Maternal blood vessel Epiblast Hypoblast Trophoblast 2 Blastocyst implants (7 days after fertilization). Figure 47.12c Expanding region of trophoblast Amniotic cavity Epiblast Hypoblast Yolk sac (from hypoblast) Extraembryonic mesoderm cells (from epiblast) Chorion (from trophoblast) 3 Extraembryonic membranes start to form (10–11 days), and gastrulation begins (13 days). Figure 47.12d Amnion Chorion Ectoderm Mesoderm Endoderm Yolk sac Extraembryonic mesoderm Allantois 4 Gastrulation has produced a three-layered embryo with four extraembryonic membranes. Developmental Adaptations of Amniotes • The colonization of land by vertebrates was made possible only after the evolution of – The shelled egg of birds and other reptiles as well as monotremes (egg-laying mammals) – The uterus of marsupial and eutherian mammals © 2011 Pearson Education, Inc. • The four extraembryonic membranes that form around the embryo – – – – The chorion functions in gas exchange The amnion encloses the amniotic fluid The yolk sac encloses the yolk The allantois disposes of waste products and contributes to gas exchange © 2011 Pearson Education, Inc. Organogenesis • During organogenesis, various regions of the germ layers develop into rudimentary organs • Early in vertebrate organogenesis, the notochord forms from mesoderm, and the neural plate forms from ectoderm © 2011 Pearson Education, Inc. Figure 47.13 Eye Neural folds Neural fold Tail bud Neural plate SEM 1 mm Neural fold Somites Neural tube Neural plate Notochord Neural crest cells 1 mm Neural crest cells Coelom Notochord Somite Ectoderm Mesoderm Endoderm Neural crest cells Outer layer of ectoderm Archenteron (a) Neural plate formation Neural tube (b) Neural tube formation Archenteron (digestive cavity) (c) Somites • The neural plate soon curves inward, forming the neural tube • The neural tube will become the central nervous system (brain and spinal cord) Video: Frog Embryo Development © 2011 Pearson Education, Inc. Figure 47.13b-3 Neural fold Neural plate Neural crest cells Neural crest cells (b) Neural tube formation Neural tube Outer layer of ectoderm Figure 47.13c Eye SEM Neural tube Notochord Coelom Somites Tail bud 1 mm Neural crest cells Somite (c) Somites Archenteron (digestive cavity) Figure 47.14 Neural tube Notochord Eye Forebrain Somite Coelom Endoderm Mesoderm Ectoderm Archenteron Lateral fold Heart Blood vessels Somites Yolk stalk These layers form extraembryonic membranes. (a) Early organogenesis Yolk sac Neural tube YOLK (b) Late organogenesis Mechanisms of Morphogenesis • Morphogenesis in animals but not plants involves movement of cells © 2011 Pearson Education, Inc. Figure 47.15-1 Ectoderm Figure 47.15-2 Ectoderm Neural plate Microtubules Figure 47.15-3 Ectoderm Neural plate Microtubules Actin filaments Figure 47.15-4 Ectoderm Neural plate Microtubules Actin filaments Figure 47.15-5 Ectoderm Neural plate Microtubules Actin filaments Neural tube • The cytoskeleton promotes elongation of the archenteron in the sea urchin embryo • This is convergent extension, the rearrangement of cells of a tissue that cause it to become narrower (converge) and longer (extend) • Convergent extension occurs in other developmental processes • The cytoskeleton also directs cell migration © 2011 Pearson Education, Inc. Figure 47.16 Programmed Cell Death • Programmed cell death is also called apoptosis • At various times during development, individual cells, sets of cells, or whole tissues stop developing and are engulfed by neighboring cells • For example, many more neurons are produced in developing embryos than will be needed • Extra neurons are removed by apoptosis © 2011 Pearson Education, Inc. Concept 47.3: Cytoplasmic determinants and inductive signals contribute to cell fate specification • Determination is the term used to describe the process by which a cell or group of cells becomes committed to a particular fate • Differentiation refers to the resulting specialization in structure and function © 2011 Pearson Education, Inc. Figure 47.17 Epidermis Central nervous system Notochord Epidermis Mesoderm Endoderm Blastula Neural tube stage (transverse section) (a) Fate map of a frog embryo 64-cell embryos Blastomeres injected with dye Larvae (b) Cell lineage analysis in a tunicate Figure 47.21 Dorsal Right Anterior Posterior Left Ventral (a) The three axes of the fully developed embryo Animal pole Animal hemisphere Vegetal hemisphere Vegetal pole (b) Establishing the axes Point of sperm nucleus entry Gray crescent Pigmented cortex Future dorsal side First cleavage Restricting Developmental Potential • Hans Spemann performed experiments to determine a cell’s developmental potential (range of structures to which it can give rise) • Embryonic fates are affected by distribution of determinants and the pattern of cleavage • The first two blastomeres of the frog embryo are totipotent (can develop into all the possible cell types) © 2011 Pearson Education, Inc. Figure 47.22-1 EXPERIMENT Control egg (dorsal view) Experimental egg (side view) 1a Control group Gray crescent 1b Experimental group Gray crescent Thread Figure 47.22-2 EXPERIMENT Control egg (dorsal view) Experimental egg (side view) 1a Control 1b Experimental group group Gray crescent Gray crescent Thread 2 RESULTS Normal Belly piece Normal Formation of the Vertebrate Limb • Inductive signals play a major role in pattern formation, development of spatial organization • The molecular cues that control pattern formation are called positional information • This information tells a cell where it is with respect to the body axes • It determines how the cell and its descendents respond to future molecular signals © 2011 Pearson Education, Inc. • The wings and legs of chicks, like all vertebrate limbs, begin as bumps of tissue called limb buds © 2011 Pearson Education, Inc. Figure 47.24 Anterior Limb bud AER ZPA Posterior Limb buds 50 m 2 Digits Apical ectodermal ridge (AER) Anterior 3 4 Ventral Proximal Distal Dorsal Posterior (a) Organizer regions (b) Wing of chick embryo Figure 47.24b 2 Digits Anterior 3 4 Ventral Distal Proximal Dorsal Posterior (b) Wing of chick embryo Figure 47.25 EXPERIMENT Anterior New ZPA Donor limb bud Host limb bud ZPA Posterior RESULTS 4 3 2 2 4 3 • Sonic hedgehog is an inductive signal for limb development • Hox genes also play roles during limb pattern formation © 2011 Pearson Education, Inc. Cilia and Cell Fate • Ciliary function is essential for proper specification of cell fate in the human embryo • Motile cilia play roles in left-right specification • Monocilia (nonmotile cilia) play roles in normal kidney development © 2011 Pearson Education, Inc. Figure 47.26 Lungs Heart Liver Spleen Stomach Large intestine Normal location of internal organs Location in situs inversus